
Most of us know what a nerve cell is, at least generally. It usually consists of a soma (body) with dendrites that collect information, and an axon that carries information about when the cell fires an action potential to other cells, often over long distances. But how and when did the neuron evolve? How did the various systems that use neurons evolve? How much and what kind of communication did the animals before nerves use, and for what?
I'm going to address these questions in this post, starting with the model I proposed earlier:
So the question is, sort of internal communication did this animal have, and what was the path followed by the lineages leading to the eumetazoan nerve cell? ... Before we address that question, however, we need to take a closer look at the animal involved, and it's evolution.Thus, we have a small ball of cells, with a bunch of collared flagella, probably in front, ciliated cells along the equator, and some sort of sensory abilities.
[...]
It's completely plausible, IMO, that a small animal like a floating sponge might have had a system of internal water-canals, while still having a regular shape and the ability to swim. Thus, each lineage would have branched off from the main line (with swimming adults), then developed a sessile lifestyle.
[...]
It may well be that there was a steady progression of advance in developmental mechanisms, with one lineage of sponges branching off after each advance. Thus, the last branch, the Homoscleromorpha, would have had almost the full suite of mechanisms, probably including biradial symmetry.
[...]
Finally, let me mention communication. It seem unlikely that this animal had nerve cells (although it's been suggested[1]), however much of the cellular machinery needed for chemical communication is present in at least some lineages of sponges.[1] [I've updated the footnotes.]
The Nature of the Urmetazoan
I've mentioned the way we can look at the paraphyly of the sponges and what it can tell us regarding the progressive evolution of the various improvements found in the eumetazoans relative to what we normally see in (adult) sponges.[2] Quoting from the referenced paper:
[...] The demonstration that Porifera is paraphyletic and therefore represents an evolutionary grade has important implications for the polarization of character states and understanding the sequence of character acquisition at the base of Metazoa.Let's take a look at what that means, given our model of a small, free-swimming spheroidal animal with internal water canals:Metazoa (Fig. 2, node 1) is characterized, in part, by the acquisition of multicellularity and the presence of the extracellular matrix, a complex of collagen, proteoglycan, adhesive glycoprotein and integrin, which mediates cell motility and transitions between epithelial and motile cell types ([ref]). Because of sponge paraphyly, the WCS [water channel system] with choanocytes (itself a likely plesiomorphic cell type), had also evolved by this point as well. The unnamed clade Calcarea + Epitheliozoa (epitheliozoans are the homoscleromorph + eumetazoans, see below) (Fig. 2, node 2) is potentially characterized by the presence of cross-striated rootlets. Most metazoan ciliated cells have a system of cross-striated rootlets that originates in the ciliary basal body and extends into the cytoplasm. Calcisponge larvae, as well as those of homoscleromorphs, have long, crossstriated cell rootlets ([refs]) that were perhaps incorporated into adult eumetazoans through neotenous evolution ([ref]). However, the choanoflagellate [arguably the closest relative to the metazoans, see here] Monosiga, the placozoan Trichoplax, as well as several other protistan taxa, also have striated rootlets ([refs]) implying that either the rootlets of Calcarea + Epitheliozoa are not homologous with those of choanoflagellates or this trait is plesiomorphic for Metazoa and has been secondarily lost in Hexactinellida and Demospongiae.
The clade Homoscleromorpha + Eumetazoa is herein recognized as Epitheliozoa (Fig. 2, node 3). Ax ([ref]) defined the clade Epitheliozoa for the clade of epithelial animals, and it is usually considered to include Ctenophora, Cnidaria and triploblasts (e.g. [ref]). The position of the homoscleromorphs as the sister taxa to Eumetazoa, as well as the presence of basal laminae ([refs]), suggests that the Epitheliozoa should include the Homoscleromorpha. A second potential apomorphy of the Epitheliozoa is the presence of an acrosome ([refs]). Thus, of the four primary eumetazoan characters—tissues, an acrosome, a nervous system and a gut—the acquisition of tissues and the acrosome antedated the last common ancestor of homoscleromorphs and eumetazoans. The expression of features such as epithelia in the adult, along with the acquisition of these new characters (nervous system and gut), and the loss of the WCS, could be due to a coordinated character change ([ref]) accompanying the neotenous evolution of a non-feeding sponge larva to a predatory eumetazoan.[2]
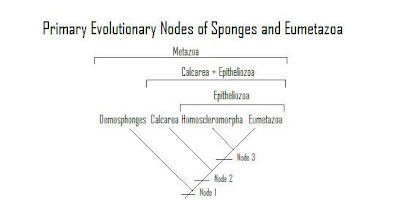
- Node 1: Metazoa:
- multicellularity
- extracellular matrix
- WCS (Water Canal System) with choanocytes
- post-synaptic protein homologs[1] (see below)
- Node 2: Calcarea + Epitheliozoa:
- cross-striated rootlets
- Node 3: Epitheliozoa:
As you can see, according to Sperling et al., the metazoa already possessed the water canal system and the extra-cellular matrix. The primary development leading to the next mode was cross-striated rootlets, although there is considerable doubt about this development. Finally, the epithelium characterizes the final node that includes modern sponges. Each of these nodes, then, presumably underwent an adaptive radiation as free-swimming animals after refining its development(s), but only a few sessile, benthic, lineages survived when the next radiation came. Only the Eumetazoans actually had free-swimming lineages that survive today.

We can imagine that the original animal, then, had refined its system of water canals, using cilia to swim and some sort of visual and olfactory (chemotaxic) system to steer, allowing it to chase down the populations of bacteria or other food they required. The development of cross-striated rootlets probably allowed the skin-mounted cilia to apply greater pressure to the water as they pushed it, permitting greater size and speed. The development of the epithelia allowed the extra-cellular area to be kept under pressure relative to the surroundings.
An important difference with Sperling et al. is that I'm assuming that the common ancestor's adult form was free-swimming, and that as each lineage evolved a sessile, benthic lifestyle, it also (independently) acquired a larval form with some vestigial characteristics of the ancestral adult.
Internal Communications
Quoting again from the model I proposed earlier:
If this model is true, our most distant ancestor might have had a fairly sophisticated system of communication long before the invention of nerve cells. It would be directly ancestral to our modern system of chemical emotions, and may have been even more sophisticated, since there were no nerve cells to handle more "digital" types of calculation. Indeed, the chemical intelligence displayed by modern vertebrates in controlling their development may well have been used to control active behavior in this ancient ancestor.Note here that the presence of a full epithelium permits isolating those chemical communications from the outside, as well as potentially compartmentalizing chemical communications into separate modules.
In three recent post, Nerve Cells and Glial Cells: Redefining the Foundation of Intelligence, Beyond the Synapse, and The Analog Axon, I discussed the large variety of inter-cell communications that don't depend on action potentials flowing along the axon. Not only nerve cells, but at least one, and likely two, types of glial cells can fire action potentials, although they may not do so in vivo. Even without action potentials, voltage variations in cell membranes can carry signals created by chemical signals from other cells. Small patches of cell membrane can act as semi-independent calculators, responding to local conditions with local releases of various transmitters. These conditions include voltage, but also the local concentrations of all sorts of signal molecules on both sides of the cell membrane.
It would be easy to assume that the sophisticated communications just mentioned evolved after the neuron did, but it's more plausible that they were there first, and that the action potential evolved later, as a high-speed, long-distance mechanism for carrying a small subset of time-dependent control information. Thus, we can assume that, by the time the ancestors of the eumetazoans diverged from those of the Homoscleromorphs, much of the system of calculation used in the modern brain was already present, at least in rudimentary form. In this regard, we may note that many of the genes used in brain communications appear to have already been present in even the early nodes.[1]
This type of calculation/communication is energy intensive, so it's quite likely that the original cells to undertake this process were ectodermal, since such cells would have had the best access to external oxygen. They may well have started by extending small processes toward other cells through the extra-cellular matrix, from which they released transmitters carrying their current message.
It is with the development of the epithelium (Node 3) that things really took off. Both the descendant lineages of this node appear to possess the full complement of developmental cell signaling and adhesion genes.[4] In addition, a very recent paper,[3] The evolution of Runx genes II. The C-terminal Groucho recruitment motif is present in both eumetazoans and homoscleromorphs but absent in a haplosclerid demosponge (by Anthony J Robertson, Claire Larroux, Bernard M Degnan and James A Coffman), demonstrates that a specific pair of genes that already existed in ancestral animals were linked in the common ancestor of node 3 but not before.
These two genes, between them, occupy a key position in controlling cell differentiation, and the fact that they are present at this node strongly suggests that the free-swimming ancestors had a complex organization of their bodies. Since the organization of internal water channels probably doesn't require this, the best hypothesis (IMO) is that is was required to organize the communication system.
For an animal a few millimeters long, this system probably didn't require the speed of the action potential, but the complex calculations possible to an organized structure of differentiated cells would have (potentially) made this a quite smart little animal. It seems very plausible to me that the primary advantage of the sophisticated system of developmental control was its ability to support the development of this powerful system of calculation and communication, long before it could gain the advantage of more complex bodies.
The Action Potential
It isn't only neurons that have sophisticated adaptions to carry action potentials, muscle cells do as well.[5] In fact, they may well have had them first, since they would need synchronized activity over their entire lengths. Most likely the signal speed of action potentials would not even be needed until there were muscles to respond to them, and muscle fibers are so large in comparison to axons that they could have extended a process to the appropriate calculating cell more cheaply than the cell could have done to them.
We can see the neural action potential evolving, then, as the specializations already present in muscle cells are adopted by nerve cells for high-speed communication.
Rather than control of muscles, I suspect that the original value of the neural action potential was to communicate overall chemical "emotional" state: the rudiments of the modern peripheral nervous system. Once the structure of the nerve cell was fully refined, it was then adopted to the control of muscles, a process that probably wasn't complete until the early bilaterians.
Robertson, A., Larroux, C., Degnan, B., & Coffman, J. (2009). The evolution of Runx genes II. The C-terminal Groucho recruitment motif is present in both eumetazoans and homoscleromorphs but absent in a haplosclerid demosponge BMC Research Notes, 2 (1) DOI: 10.1186/1756-0500-2-59
Links: (Some of these are duplicates of links from earlier posts. Not all have been called out in the text. Many are derived from the main article. Use the back key if you came via clicking a footnote.) (I've included only the link referenced in this leader.)
1. A Post-Synaptic Scaffold at the Origin of the Animal Kingdom
2. Poriferan paraphyly and its implications for Precambrian palaeobiology
3. The evolution of Runx genes II. The C-terminal Groucho recruitment motif is present in both eumetazoans and homoscleromorphs but absent in a haplosclerid demosponge
4. Early evolution of animal cell signaling and adhesion genes
5. t-tubules and sarcoplasmic reticulum function in cardiac ventricular myocytes
I must say this is good post since I have come acroos too many nurvous system article and as comparing this with it, It is good in both graphics and text.
ReplyDeletedisque dur externe multimedia
This is a very interesting topic, by the way, I think your blog is very informative about this issues so congratulations for the well done your you have manage to build.
ReplyDeleteHello Dude,
ReplyDeleteReally it is a nice blog. The nervous system is an organ system containing a network of specialized cells that coordinate the actions of an animal and transmit signals between different parts of its body. These are found in most multicellular animals, but vary greatly in complexity. Thanks a lot for sharing with us...