

Find it here, unless you already did. (It's been there since the 19th.)
I kept a close watch for the first few days, then started to slack off. It was originally supposed to be hosted by Greg Laden at Quiche Moraine, and that was where I looked. Even after the announcement (on the 18th) that it would be published "tomorrow", I looked there. And didn't find it, and didn't find it, etc. It just never occurred to me to look back at Living the Scientific Life (Scientist, Interrupted) for updates, where I would have found the post itself.
If anybody was waiting for me to announce Scientia Pro Publica, sorry. But I suspect anybody really interested would have been Googling it, and found it before I did. But if you aren't interested, you should be. Lots of good stuff there.
Read more!
Saturday, August 29, 2009
Thursday, August 20, 2009
Semantic Strait-Jackets in Science
While working on an earlier post, I ran into an annoying issue: in synapses, the result of a surge of GABA or Glycine is called and IPSP, meaning Inhibitory Post Synaptic Potential. This remains true even when the effect is actually excitatory. This is annoying, but easy enough to live with (for me) as I tend to think in historic terms, especially with regard to words.
But it brought to mind an interesting point: science is full of terms and definitions that are obsolete, with research subsequent to naming and definition having rendered the specific names, and/or some of the assumptions that went into them, invalid.
The case in point makes a good illustration: the assumption that a particular synaptic action in response to an action potential is inhibitory is built into the name, and it takes an effort of will and memory to keep track of the various times that it might be (or is known to be) excitatory.
But wait! There's more...
Not only does this semantic confusion result in lost time and errors when somebody forgets to allow for all the naming exceptions, but it also tends to channel everybody's thoughts into an assumption that the result of an action potential at a synapse is either one or the other, and that's it.
The reality, especially with chloride currents, is much more complex. ... Even if a burst of GABA causes the membrane to depolarize, the equilibrium voltage (also called the "reversal potential") may be somewhere between the resting voltage and the action potential threshold.

Figure 1: Effect of Cl- current with Equilibrium voltage at ~-62mV as an example. (Original. You may link to, copy, and or modify this image.)
In figure 1, we see just this situation. Compare with those from Axons and Chloride Currents, where the illustrated equilibrium voltages are either below the resting voltage (figure 2), or above the action potential threshold (figure 3).

Figure 2: Effect of Cl- current with Equilibrium voltage at ~-73mV as typical for a pyramidal cell soma. (From Axons and Chloride Currents. You may link to, copy, and or modify this image.)
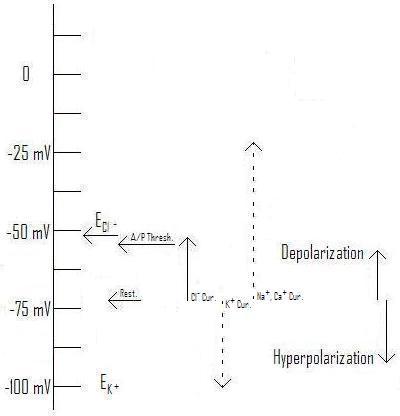
Figure 3: Effect of Cl- current with Equilibrium voltage at ~-54mV as typical for a pyramidal cell AIS. (From Axons and Chloride Currents. ; You may link to, copy, and or modify this image.)
In figure 1, a strong Cl- current will tend to depolarize the membrane, but only so far. If an EPSP based on sodium or calcium attempts to depolarize it further, the Cl- current will have a shunting action that will tend to resist this depolarization. How the membrane responds, and whether an action potential is fired, will be a very complex problem.
The very name IPSP, then, tends to prejudice our minds, hiding the complexity of what's going on. Ideally, IMO, this current should be renamed "CPSP" for "Chloride Post Synaptic Potential". This name is neutral with respect to whether it will tend to depolarize or hyperpolarize the membrane, as well as calling attention to the fact that the result is more complex than simply that binary choice.
Of course, I doubt it's going to change soon. Big Science has become as institutionalized as the Roman Catholic Church was in Galileo's day, if not as centralized in its power, and pushing such a change through would probably take as much effort as redefining Pluto as not a planet. Rather, this particular semantic strait-jacket will continue to inconvenience and distract people who haven't learned to live with it, along with all the other obsolete terms and definitions of this type.
The biggest problem I have with this isn't the distraction and tendency to hide important details. Rather, there are two ways to approach names, in science and anywhere else: you can remember them in functional terms, linking the words of the name to their everyday meanings, or you can remember them as arbitrary symbols, strings of words that represent a particular thing completely independently of their everyday meanings. When the string of words that makes up a scientific name doesn't represent the same thing as what the name is used for, only the latter way of remembering is available (unless you want to load up your brain with a bunch of the history of science). Thus, IPSP doesn't stand for an actual hyperpolarization of the membrane (which is what the words actually mean), but for a current carried by chloride or potassium ions, regardless of its effect on membrane voltage.
This turns learning about science into a process of memorizing pointless and arbitrary names, that can't be deciphered in terms of the everyday meanings of the words that make them up. Not only does this interfere with the education and development of people who would make good scientists, but it makes the field more attractive to the sort of people who don't do creative thinking: all they want to do is memorize formulas and rituals, and spend their career following the rituals they learned in college. Such people don't, IMO, really have much to contribute to science.
Read more!
But it brought to mind an interesting point: science is full of terms and definitions that are obsolete, with research subsequent to naming and definition having rendered the specific names, and/or some of the assumptions that went into them, invalid.
The case in point makes a good illustration: the assumption that a particular synaptic action in response to an action potential is inhibitory is built into the name, and it takes an effort of will and memory to keep track of the various times that it might be (or is known to be) excitatory.
But wait! There's more...
Not only does this semantic confusion result in lost time and errors when somebody forgets to allow for all the naming exceptions, but it also tends to channel everybody's thoughts into an assumption that the result of an action potential at a synapse is either one or the other, and that's it.
The reality, especially with chloride currents, is much more complex. ... Even if a burst of GABA causes the membrane to depolarize, the equilibrium voltage (also called the "reversal potential") may be somewhere between the resting voltage and the action potential threshold.
Figure 1: Effect of Cl- current with Equilibrium voltage at ~-62mV as an example. (Original. You may link to, copy, and or modify this image.)
In figure 1, we see just this situation. Compare with those from Axons and Chloride Currents, where the illustrated equilibrium voltages are either below the resting voltage (figure 2), or above the action potential threshold (figure 3).
Figure 2: Effect of Cl- current with Equilibrium voltage at ~-73mV as typical for a pyramidal cell soma. (From Axons and Chloride Currents. You may link to, copy, and or modify this image.)
Figure 3: Effect of Cl- current with Equilibrium voltage at ~-54mV as typical for a pyramidal cell AIS. (From Axons and Chloride Currents. ; You may link to, copy, and or modify this image.)
In figure 1, a strong Cl- current will tend to depolarize the membrane, but only so far. If an EPSP based on sodium or calcium attempts to depolarize it further, the Cl- current will have a shunting action that will tend to resist this depolarization. How the membrane responds, and whether an action potential is fired, will be a very complex problem.
The very name IPSP, then, tends to prejudice our minds, hiding the complexity of what's going on. Ideally, IMO, this current should be renamed "CPSP" for "Chloride Post Synaptic Potential". This name is neutral with respect to whether it will tend to depolarize or hyperpolarize the membrane, as well as calling attention to the fact that the result is more complex than simply that binary choice.
Of course, I doubt it's going to change soon. Big Science has become as institutionalized as the Roman Catholic Church was in Galileo's day, if not as centralized in its power, and pushing such a change through would probably take as much effort as redefining Pluto as not a planet. Rather, this particular semantic strait-jacket will continue to inconvenience and distract people who haven't learned to live with it, along with all the other obsolete terms and definitions of this type.
The biggest problem I have with this isn't the distraction and tendency to hide important details. Rather, there are two ways to approach names, in science and anywhere else: you can remember them in functional terms, linking the words of the name to their everyday meanings, or you can remember them as arbitrary symbols, strings of words that represent a particular thing completely independently of their everyday meanings. When the string of words that makes up a scientific name doesn't represent the same thing as what the name is used for, only the latter way of remembering is available (unless you want to load up your brain with a bunch of the history of science). Thus, IPSP doesn't stand for an actual hyperpolarization of the membrane (which is what the words actually mean), but for a current carried by chloride or potassium ions, regardless of its effect on membrane voltage.
This turns learning about science into a process of memorizing pointless and arbitrary names, that can't be deciphered in terms of the everyday meanings of the words that make them up. Not only does this interfere with the education and development of people who would make good scientists, but it makes the field more attractive to the sort of people who don't do creative thinking: all they want to do is memorize formulas and rituals, and spend their career following the rituals they learned in college. Such people don't, IMO, really have much to contribute to science.
Read more!
Axons and Chloride Currents

It's commonly assumed that synapses using GABA and glycine as neurotransmitters are inhibitory, that is when an action potential causes the release of (one or both of) these neurotransmitters (from the pre-synaptic side of the synapse) the result is a reduced chance that the post-synaptic cell will fire an action potential. However, this is not always true, there are several common circumstances under which these neurotransmitters appear to increase the chance of an action potential firing in the post-synaptic cell, as discussed in two recent papers to be discussed here. First, let's look at the mechanisms responsible for this reversal of the standard expectations, then I'll go over the two papers, then integrate the subject with some of my previous discussions regarding the transmission of analog data (besides action potential timing) along the first few hundred microns of the axon.
The role of Chloride Currents in Neural Communication
In a recent post I described the general relationship of various currents, ion channels, and voltage events (e.g. spikes and action potentials) in the neural cell membrane, but deferred discussion of chloride (Cl-) currents because of their complexity.
The key item regarding any ion-carried current is the equilibrium voltage (also often called the reversal potential): that voltage (across the cell membrane) at which the electrical pressure driving a specific ion in one direction exactly balances the concentration-driven pressure in the other. The equilibrium voltage is primarily determined by the relative concentrations on either side of the cell membrane, so a change to either concentration can produce a change to the equilibrium voltage, and different concentrations across different parts of the cell (membrane) will result in different equilibrium voltages. (They will also result in a net ion diffusion, along with an energy cost to maintain it: see below.)

Figure 1: Action Potential Threshold and Directions of Depolarization and Hyperpolarization Relative to Figure 3 of A New Integrative Theory for Cortical Pyramidal Neurons. (From Figure 4 of the same post.)
As you can see from figure 1, the typical equilibrium voltage for Cl- falls right into the range of typical "resting" voltages across the membrane. All of these are within about 40 mV of a typical action potential threshold, which is primarily determined by the nature of the specific voltage-gated Sodium (Na+) channels present in that piece of membrane. (See my discussion here for more detail.) Now, let's compare the effects of changing the Cl- concentration inside the cell from that of the soma (body) of a typical pyramidal cell to that of the axon initial segment[A1] (AIS: the first part of the axon after it leaves the axon hillock).
Figure 2: Effect of Cl- current with Equilibrium voltage at ~-73mV as typical for a pyramidal cell soma.[1] (Original. You may link to, copy, and or modify this image.)
Figure 3: Effect of Cl- current with Equilibrium voltage at ~-54mV as typical for a pyramidal cell AIS.[1] (Original. You may link to, copy, and or modify this image.)
As you can see, raising the equilibrium voltage for Cl- changes the direction of the current and its effect. Rather than being inhibitory (hyperpolarizing), the chloride current is excitatory (depolarizing), tending to make the post-synaptic cell more likely to fire an action potential. Notice in this illustration (Figure 3) the equilibrium voltage is higher than the threshold for firing an action potential, so enough chloride current can "do the job" independently of help from depolarizing influences coming from the dendrites. This has actually been observed in experiments.[1] [21]
As mentioned in A New Integrative Theory for Cortical Pyramidal Neurons, the normal concentration difference is just about enough to create an equilibrium voltage in the same range as the typical resting potential. This is because the concentration of Cl- outside the cell is greater than that inside by just enough to counteract the resting voltage. If, somehow, a higher concentration of Cl- is present on the inside of the membrane, the equilibrium voltage will rise, as shown in Figure 3 for the AIS.
Now, we can't just leave it that the inhibitory effects of neurotransmitters such as GABA and glycine can be reversed, we need to consider both the receptors (all types) and and the effects of changing the voltage under inhibitory conditions. ... (read the rest in the full post)
Let's start with Figure 2, when a big dose of GABA arrives at a synapse. The fastest-acting receptor(s) for GABA are the GABAA receptors, in which the molecule that spans the membrane contains both a pore through which the Cl- ions can pass, and a receptor region (outside the cell) that fits to GABA like a lock to a key, changing its shape (and the shape of the molecule), and allowing the pore to open. (A good discussion of this receptor may be found at The Versatile GABAa Chloride Channel Receptor Complex at Physiology physics woven fine. I'm not going to try duplicate this work, but in reading it just keep in mind that the author has not considered that the GABAA receptor can sometimes be excitatory.)
Notice, though, that the difference in voltages is very small. This isn't as important is it might seem, because when the depolarizing activity of an excitatory synapse (a regular one using i.e. glutamate or acetylcholine) tries to raise (depolarize) the voltage towards the threshold, the greater difference in voltage drives a greater current, tending to prevent the voltage from going very far from the resting potential.
By contrast, a potassium (K+) current will pull the voltage much farther down, distinctly hyperpolarizing the cell membrane, not just at the synapse but a considerable distance away. This will become important later (below), because another GABA receptor, the GABAB receptor, can cause K+ channels to open, having just this effect. Individual cells can express different mixes of receptors, and even the same cell can have different mixes in different parts of the cell.[27] [28]
This isn't just true of receptors, either. Ion pumps can be localized, with massively different densities in different parts of the cell membrane (as can many other membrane proteins.[27]
Now, let's look at the AIS, in Figure 3. Here, a big dose of GABA opens the pores in the GABAA receptor in the same way as above, but it produces a distinct depolarizing current, pulling the membrane voltage, as always, towards its equilibrium voltage, which in this case actually happens to be higher than the action potential threshold. Thus, in this case the GABA can not only be excitatory, it can trigger an action potential. (Of course, if the post-synaptic cell also expresses GABAB receptors, the effect will be much more complex: see below.)
Chloride Concentrations and Their Maintenance
What are the typical CL- concentrations driving these different currents? I'm not going to go into the extra-cellular (concentration outside the cell) concentration, since they're the same for all parts of the cell. (Since different parts of different cells are bathed in the same local extra-cellular fluids.) Instead we'll look at the intra-cellular (concentration inside the cell) found by Khirug, Yamada et al., one of the papers we'll be discussing.[20]
By determining the reversal potentials mediated by sudden release of GABA("EGABA values of –59.4 ± 1.5 mV (n = 14), –65.8 ± 1.2 mV (n = 14), and –70.9 ± 1.5 mV (n = 10), respectively (Fig. 1)"[20]), they could calculate the concentrations:
Assuming an intracellular pH of 7.2, the intracellular levels of Cl– ([Cli]) calculated on the basis of the above EGABA values ([refs]) are 11, 7.9, and 6.0 mM [milliMoles, a measure of concentration] for the AIS, soma, and dendrite, respectively.[20]Of course these values are typical of only one type of cell (dentate gyrus cells (DGCs) of the hippocampus of mice and rats), but they give us some numbers to work with.
How are these concentrations maintained against the inevitable leaks and currents, which would normally push ECl- towards the resting voltage?[20] [33] There are two specific transporters involved (out of a large number used throughout the body): Both are members of a large family called cation–chloride co-transporters (CCCs), transmembrane proteins that transport a specific mix of ions in a compulsory group (all together or none). One is a potassium-chloride co-transporter (KCC): KCC2, while the other is a sodium-potassium-chloride co-transporter (NKCC): NKCC1. (The numbers are confusing, the "1" has nothing to do with the "2": the body also has KCC1 and NKCC2, although they aren't relevant to this discussion.)
KCCs, including KCC2, transport one K+ and one Cl- together, which is plenty to maintain the type of potential seen in Figure 2. The K+ has a strong pressure "pushing" it out of the cell, and it will drag the Cl- along with it. (Remember that since these ions have opposite charges, they represent currents going in opposite directions.)
NKCCs, including NKCC1, move one Na+, one K+ and two Cl- together, which means they will go in the other direction because the very high pressure "pulling" Na+ into the cell will trump the pressure of the other three ions.
During development, most neurons express NKCC1, making GABAA currents excitatory, which appears to be necessary for proper neural development.[2] [20] Mature neurons appear to mostly express KCC2 in the soma and dendrites, which makes Cl- currents hyperpolarizing. There has been some controversy over how the higher concentrations in the AIS were maintained, with the standard explanation being the absence of KCC2 in the AIS.
GABAergic Depolarization of the Axon Initial Segment in Cortical Principal Neurons Is Caused by the Na–K–2Cl Cotransporter NKCC1
This paper, (by Stanislav Khirug, Junko Yamada, Ramil Afzalov, Juha Voipio, Leonard Khiroug, and Kai Kaila,) investigates the conflict between the point (noted above) that the inevitable leaks and currents would normally push ECl- towards the resting voltage, and the fact that Cl- currents in the AIS have been observed to be distinctly depolarizing, which means that the concentration must be maintained at a level such that ECl- is distinctly higher than the resting voltage. A simple absence of KCC2, which would push ECl- lower, wouldn't cut it.
First, by subjecting different parts of the same cell to bursts of GABA (with any GABAB receptors blocked using "CGP55845A [(2S)-3-[(15)-1-(3,4-dichlorophenyl)ethyl]amino-2-hydroxypropyl)-(phenylmethyl)phosphinic acid]"[20]) and measuring the resulting equilibrium voltages, they demonstrated the different values of EGABA for the AIS, soma, and dendrites. This may reasonably be assumed to be close to the value for ECl-, with only the GABAA receptors working.
Then, they did identical experiments similar to the one described above with both wild type mice (WT) and mutants unable to express NKCC1 anywhere(NKCC1–/–). The results:
The WT neurons showed an axo-somatic ΔEGABA that was similar (5 mV) to the one described above in the Thy1–mGFP neurons with AIS, somatic, and dendritic EGABA values of –61.3 ± 2.4 mV (n = 14), –66.0 ± 1.6 mV (n = 22), and –71.0 ± 2.0 mV (n = 16), respectively. Notably, in identical recordings from the NKCC1–/– neurons, there was no axo-somatic EGABA gradient; the values for EGABA at the AIS and soma were similar, –70.5 ± 1.5 mV (n = 17) and –70.3 ± 1.5 mV (n = 18), respectively. However, a somato-dendritic ΔEGABA similar to that in the WT neurons was still observed (dendritic EGABA of –74.6 ± 1.7 mV; n = 15). These data indicate a key role for NKCC1 in the generation and maintenance of the chloride gradient that results in depolarizing GABA responses in the AIS of mouse DGCs and suggest that NKCC1 has no significant influence on dendritic EGABA. Interestingly, the above observations are not compatible with the idea that the somato-dendritic EGABA gradient is set by NKCC1 and KCC2. The putative transport mechanism underlying this gradient is not in the focus of the present work, but preliminary observations point to a role of a bicarbonate-dependent somatic transporter that accumulates chloride in the DGCs (our unpublished data). [20]They have thus demonstrated, for two types of cells, the DGCs and "rat neocortical layer 2/3 pyramidal neurons", that the excitatory (depolarizing) action of GABA synapses onto the AIS is probably caused by the expression and localized targeting of NKCC1. They've also discovered another effect evidently not caused by NKCC1: the difference in Cl- concentrations between the soma and dendrites. It'll be interesting to see what further research turns up in this direction.
Complex Events Initiated by Individual Spikes in the Human Cerebral Cortex
This paper (by Gábor Molnár, Szabolcs Oláh, Gergely Komlósi, Miklós Füle, János Szabadics, Csaba Varga, Pál Barzó, Gábor Tamás) reports intriguing results from "complex events triggered by individual action potentials in the human neocortical network." Unlike previous research, they used slices of human brain with part of the small-scale network intact, and applied single strong spikes to a single cell and observed the result.
Before I go on with this, let me save a lot of readers the effort of digging into the paper over ethical concerns with this blockquote:
All procedures were performed according to the Declaration of Helsinki with the approval of the University of Szeged Ethical Committee. Human slices were derived from material that had to be removed to gain access for the surgical treatment of deep-brain tumors from the left and right frontal, temporal, and parietal regions with written informed consent of the patients (aged 18–73 y) prior to surgery over the last 4 y.[21](Anybody wanting more information can use part of the above blockquote in a text search in the article, which is open access.)
I'm not going to spend further on the methods, or the details of the results, instead jumping to a more "high-level" discussion, starting with another blockquote:
Our results show that a single spike of a pyramidal cell in the human cortical microcircuit is capable of activating complex sequences of postsynaptic potentials lasting an order of magnitude longer than detected previously [refs]. The initiation and internally precise temporal structure of these event series appears to follow a stereotyped mechanism of spike-to-spike transmission traveling through a subset of synaptically connected neurons that seems to be conserved across several brain regions of the human cerebral cortex. The flow of downstream activation that follows the first-order spike of the trigger pyramidal cell is directionally controlled at two consecutive synaptic steps. Second-order spikes are triggered exclusively in GABAergic interneurons and not in pyramidal cells due to interneuron-selective EPSPs of enormous amplitude. In turn, second-order spikes in axo-axonic cells give rise to third-order spikes detected only in pyramidal cells, resulting in trisynaptic EPSPs in the network because axo-axonic cells do not innervate other GABAergic cells [refs]. Synchronized to the spikes in axo-axonic cells, second-order spikes in basket cells and possibly in other types of interneuron [refs] elicit hyperpolarizing effects reported here as disynaptic IPSPs.What does this mean? The human brain, (or at least the parts of the neocortex investigated,) is a very complex network of cells with positive (excitatory) and negative (inhibitory) effects, in which a single spike applied to a resting network ends up producing a stereotyped mix of these effects. Basically, there is a "cascade" (a bad analogy, but...) of action potentials, starting with the artificially triggered
- "first-order spike", followed by one or more
- "Second-order spikes" in "GABAergic interneurons", including axo-axonic cells which have excitatory GABAergic synapses on the AISs of pyramidal cells, as well as "basket cells and possibly in other types of interneuron" which have inhibitory GABAergic synapses on the somas and dendrites of pyramidal cells, followed in turn by
- "third-order spikes detected only in pyramidal cells, resulting in trisynaptic EPSPs [excitatory postsynaptic potentials] in the network",
One of the key points, for our purposes, is that the "axo-axonic cells do not innervate other GABAergic cells", they terminate only on the AIS of pyramidal cells.[34] This means that the very excitatory GABAergic synapses we've been discussing play an essential role in this network process.
Beyond Action Potentials
So far, we've covered the currents resulting from action potentials in GABAergic synapses terminating on the AIS. But regular readers here will probably recall The Analog Axon, where I concluded:
It's not a totally new discovery, of course, that some nerve cells use more analog types of communication than just action potentials.[refs] Sensory nerves especially, often make use of analog calculations. Nevertheless, except for the sensory margins of cognition, the central nervous system is usually pictured as communicating through action potentials that carry only their relative timing as information. Over long distances, this is probably true.This is particularly important for the pyrmidal cells involved in the "third-order spikes" mentioned above, where the axo-axonic input has set up varying chloride currents--varying both along the AIS and over time. These currents set the membrane voltage, often to a point substantially above (more depolarizing) the nominal "resting" voltage, with equally varying effects on the size, length, and shape of the action potential, in addition to their contribution to the probability and timing of firing the action potential in the first place. While over longer distances these variations will go away, short collaterals to nearby neurons will cary them. Studies on the fixing the membrane voltage at the soma have shown effects propagating over 400 microns (μ),[15], and we can probably add the length of the AIS to similar effects generated by voltage changes there. Similar distances for propagating analog effects have been found in presynaptic hippocampal mossy fiber boutons,[14] the output synapses from mossy fiber cells where:However, a great many of the connections in the brain, including the neocortex, arguably the most important part of the relative expansion of the human brain, are within a few hundred microns, the distance discussed here.
Excitatory presynaptic potentials result from subthreshold dendritic synaptic inputs, which propagate several hundreds of micrometers along the axon and modulate action potential–evoked transmitter release at the mossy fiber–CA3 synapse.[14]Note that these effects in the dendrites have to cross the soma before they can extend "several hundreds of micrometers along the axon", presumably similar subthreshold synaptic inputs landing on the AIS would propagate farther.
How many synapses are we talking about to be affected by these inputs? From the research mentioned above fixing the membrane voltage of the soma:[15]
Layer 5 pyramidal neurons, like other cortical neurons, give rise to a local high density of axonal connections to other pyramidal and non-pyramidal cells[refs] (Supplementary Figs 1, 4 and 7). Indeed, examination of the main axon and local axon collaterals of biocytin-filled layer 5 pyramidal cells from our slices revealed, on average, 155 (±79; n=14 cells) putative synaptic boutons within 0.5-mm of the cell body, and 269 (±152) putative synaptic boutons within the first 1mm(Fig. 4c and Supplementary Fig. 7). These values are probably a significant underestimate of local synaptic connectivity, owing to the cutting of axons and limitations of axonal staining using the slice technique (see Supplementary Figs 1 and 4).[15]This means that the AIS and short collaterals of each pyramidal cell can potentially act as an independent integrator of axo-axonic inputs, feeding a real-time analog output from hundreds of inputs[31] to hundreds of outputs[15] through the size, duration, and shape of action potentials, as well as sub-threshold changes that could affect the size of the transmitter release burst at the synapse. Moreover, these effects are tunable: addition of GABAB receptors to a synapse could produce a longer-term (50-300 ms) K+ current counteracting the depolarizing effect when many bursts of GABA arrive within a short time.
Like many other recent discoveries, these add yet more complexity, and potential calculating power, to the overall activity of the brain. This is an effect that will be very difficult to model in the more traditional neural networks, hammering home yet again how we must not limit our ideas of the brain's ability by what current models and neural networks can do. There's more where that came from, and the next decade will almost certainly see increasingly complex and powerful models of what the brain can do.
Khirug, S., Yamada, J., Afzalov, R., Voipio, J., Khiroug, L., & Kaila, K. (2008). GABAergic Depolarization of the Axon Initial Segment in Cortical Principal Neurons Is Caused by the Na-K-2Cl Cotransporter NKCC1 Journal of Neuroscience, 28 (18), 4635-4639 DOI: 10.1523/JNEUROSCI.0908-08.2008
Molnár, G., Oláh, S., Komlósi, G., Füle, M., Szabadics, J., Varga, C., Barzó, P., & Tamás, G. (2008). Complex Events Initiated by Individual Spikes in the Human Cerebral Cortex PLoS Biology, 6 (9) DOI: 10.1371/journal.pbio.0060222
Appendix: You can use the back key to return to where you were, or click on the quoted text to return to the line with the footnote.
A1. "axon initial segment" There's no link to Wiki because, as of this writing, I couldn't find anything on Wiki discussing it. So herewith...
As mentioned above, the AIS is the first part of the axon, "from the narrow beginning of the axon to the onset of the myelin sheath" (often more than 200 microns (μ))[35], or in unmyelinated axons "a variable distance, 4-5 μ." It has recognizable features:[31]
The initial segment of the axon is unlike any other known process of the nerve cell, and in certain respects it is unlike any other part of the axon itself. In the idealized nerve cell it arises from the summit of a conical projection on the surface of the perikaryon, the axon hillock (Fig. 1). The surface of the axon hillock is bounded by a plasma membrane with the usual trilaminate structure, but near the apex of the hillock (Fig. 2) a thin, dark layer of finely granular material appears just beneath the membrane. In low-power electron micrographs this granular material gives the membrane a dense appearance, so that on superficial examination the plasmalemma seems to be reduplicated or thickened. But the undercoating is not really a part of the limiting membrane; instead, it is a thin layer of powdery densities, about 100 Å [Ångströms] thick, separated from the surface membrane by a clear interval of about 30 Å (Figs. 3 and 5). As its margins are not distinct, these measurements cannot be precise. The undercoating extends from the narrow beginning of the axon to the onset of the myelin sheath, where it ceases as abruptly as it began. In unmyelinated axons the undercoating extends for a variable distance, 4-5 μ. Where the axon originates from a dendrite, the undercoating has the same sudden onset and is accompanied by other changes (see below) in the internal structure of the process that signal the beginning of an axon.[31]The above text, and the figures it references, are in Reference 31, which is open access, although it's rather old.
Both the axon hillock and the AIS are distinct compartments within the cell, which can be independently targeted by distinct mixes of specific receptors, ion pumps, and all the other types of machinery the cell uses.[27] [28] [29] Ultrastructurally:
In the axon hillock (Fig. 2) microtubules collect into bundles of three to five or more, which funnel into the initial segment and run parallel with one another throughout its length. Large initial segments have five or six such fascicles, while small ones may have only one. The number of microtubules in a bundle varies considerably but is probably characteristic of the type of neuron. In the Deiters cells of the lateral vestibular nucleus (Fig. 5) and in the motor neurons of the spinal cord, the number of microtubules included in a fascicle is usually three to five, and in the pyramidal cells of the cerebral cortex (Fig. 6) the number can reach 22. In the Purkinje cell of the frog, Kohno (13) found from 6 to 25 microtubules in a bundle, but in the rat the maximumThese "synaptic boutons attached to the surface of the perikaryon at the axon hillock" and "the initial segment of the cerebral pyramidal cell" are at the center of the primary thrust of this post.
number we have found in this type of neuron is only 10-12.In longitudinal sections the fasciculated microtubules in the initial segment often appear darker than the single microtubules in the rest of the nerve cell and its processes. This appearance is due only partly to overlapping of the microtubules in a bundle within the thickness of the section. In addition, each microtubule is surrounded by a cloud of fine fibrillar material that contributes to the general density of the fascicle. In transverse sections it can be seen that the microtubules are arrayed close together in a curving and sometimes branching line (Figs. 3, 4, and 6). Single or isolated microtubules are rarely encountered in the initial segment. Favorably oriented transverse sections show that the microtubules within the fascicles are bound together by thin, dark crossbars or arms (Figs. 3 and 6).
The bundling of the microtubules ceases abruptly at the beginning of the myelin sheath. Whether they continue down the axons as isolated microtubules or are replaced by new tubules beginning in this region could not be determined from the sections that we examined.
Although the axon hillock and the beginning of the axon fail to stain with basic dyes, clusters and rosettes of ribosomes do occur in the axon hillock and, in diminishing quantities, throughout the length of the initial segment. Apparently they are not numerous or concentrated enough to produce a basophilia that is recognizable in the light microscope. The ribosomes are usually, but not always, associated with a tubule or two of the endoplasmic reticulum. At the beginning of the myelin sheath they disappear while the endoplasmic reticulum continues in its agranular form throughout the axon.
Other cytoplasmic components of the axon, the neurofilaments, the mitochondria, multivesicular bodies, and various vesicles, all pass into the axon from the axon hillock without undergoing any distinctive change in their appearance or aggregation. Near the apex of the axon hillock the mitochondria, neurofilaments, microtubules, and endoplasmic reticulum all assume a remarkably parallel orientation as they funnel into the narrow initial segment.
It is common to find synaptic boutons attached to the surface of the perikaryon at the axon hillock, but they are unusual on the surface of most initial segments. For example, in sections through the initial segments of some 60 different Purkinje cells only one synapsing bouton was found. In contrast, nearly every section through the initial segment of the cerebral pyramidal cell shows an attached bouton (Fig. 7). No examples have yet been encountered of initial segments studded with boutons like the dendrites and perikaryon of certain cells.
In a few cases in which the apposition between an axon terminal and the initial segment was caught in a favorable plane of section, it was possible to see that the typical undercoating of finely granular material was interrupted at such sites and that the surface of the axon reverted to the appearance it normally has in the internodal segments. Only at the location of the "synaptic complex" or "active zone" was there a deviation from the normal, here resulting from the aggregation of fine filamentous material that formed the postsynaptic density. The typical undercoating of the initial segment resumed beyond the margin of the apposing terminal. If a neuroglial process was inserted between the terminal and the axon, breaking the apposition, the typical undercoating reappeared beneath it. In our material the number of cases in which we could clearly follow both the pre- and postsynaptic membranes was too small to allow us to generalize this description with assurance.[31]
The AIS is the most common site for an action potential to begin, followed by the axon hillock. As the general dendritic excitement (depolarization) increases, the site often moves back, first to the soma, then sometimes to the proximal dendrite(s).[35]
Links: (I've only included the links called out in this leader.) Not all of these are called out in the text. Use the back key if you came via clicking a footnote above.
1. Excitatory Effect of GABAergic Axo-Axonic Cells in Cortical Microcircuits requires free registration
2. Cation–chloride co-transporters in neuronal communication, development and trauma paywall
3. Chloride is preferentially accumulated in a subpopulation of dendrites and periglomerular cells of the main olfactory bulb in adult rats paywall
4. Cluster Analysis–Based Physiological Classification and Morphological Properties of Inhibitory Neurons in Layers 2–3 of Monkey Dorsolateral Prefrontal Cortex Open Access
5. Output of neurogliaform cells to various neuron types in the human and rat cerebral cortex Open Access
6. A Population of Prenatally Generated Cells in the Rat Paleocortex Maintains an Immature Neuronal Phenotype into Adulthood paywall Is this one relevant?
7. A physiological role for GABAB receptors and the effects of baclofen in the mammalian central nervous system paywall
8. A subset of periglomerular neurons in the rat accessory olfactory bulb may be excited by GABA through a Na+-dependent mechanism paywall
9. Channel behavior in a gamma-aminobutyrate transporter Open Access
10. Excitatory amino acid transporter 5, a retinal glutamate transporter coupled to a chloride conductance Open Access
11. Structure of intraglomerular dendritic tufts of mitral cells and their contacts with olfactory nerve terminals and calbindin-immunoreactive type 2 periglomerular neurons paywall
12. Compensatory changes in cellular excitability, not synaptic scaling, contribute to homeostatic recovery of embryonic network activity paywall
13. Enigmatic Central Canal Contacting Cells: Immature Neurons in "Standby Mode"? paywall
14. Combined Analog and Action Potential Coding in Hippocampal Mossy Fibers Requires free registration
15. Modulation of intracortical synaptic potentials by presynaptic somatic membrane potential
16. Long-Term Activity-Dependent Plasticity of Action Potential Propagation Delay and Amplitude in Cortical Networks Open Access
17. Information processing by graded-potential transmission through tonically active synapses
18. Enhancement of presynaptic neuronal excitability by correlated presynaptic and post-synaptic spiking
19. Recurrent excitation in neocortical circuits paywall
20. GABAergic Depolarization of the Axon Initial Segment in Cortical Principal Neurons Is Caused by the Na–K–2Cl Cotransporter NKCC1 Open Access
21. Complex Events Initiated by Individual Spikes in the Human Cerebral Cortex Open Access
22. Axonal GABAA receptors paywall
23. Robust Short-Latency Perisomatic Inhibition onto Neocortical Pyramidal Cells Detected by Laser-Scanning Photostimulation paywall
24. Excitatory GABAergic Activation of Cortical Dividing Glial Cells paywall
25. GABA Transporter GAT1 Prevents Spillover at Proximal and Distal GABA Synapses Onto Primate Prefrontal Cortex Neurons paywall
26. GABA: a pioneer transmitter that excites immature neurons and generates primitive oscillations
27. Compartmentalizing the neuronal plasma membrane from axon initial segments to synapses paywall
28. Localization and Targeting of Voltage-Dependent Ion Channels in Mammalian Central Neurons paywall
29. The distribution and targeting of neuronal voltage-gated ion channels
30. Excitatory effects of GABA in established brain networks paywall
31. The axon hillock and the initial segment Open Access
32. Proximity of excitatory and inhibitory axon terminals adjacent to pyramidal cell bodies provides a putative basis for nonsynaptic interactions paywall
33. The cellular, molecular and ionic basis of GABAA receptor signalling
34. Salient features of synaptic organisation in the cerebral cortex
35. From molecules to networks : an introduction to cellular and molecular neuroscience edited by John H. Byrne, James Lewis Roberts, chapter 17
Read more!
Monday, August 17, 2009
Diagonal Postures & The Descent from Human to Ape
Homeotics, Cladistics and the Triple Emergence of Closed Hand Gaits in Descendants of an Upright Ancestor of the Apes
Guest Post by Aaron Filler, MD, PhD, FRCSIf an upright bipedal ancestor evolved first, why would three of its descendant lineages abandon orthograde posture and restore quadrupedal gaits to their repertoire?
Firstly, we don't have to have an answer as to why it happened if that is indeed what happened. If it occurred, then it occurred whether or not we have proposed and accepted a post-hoc adaptationist explanation for it.
This week in PNAS, a paper from Tracy Kivell and Daniel Schmitt,[9] reports on differences between the functional anatomy of knuckle walking of gorillas as distinct from the basis in chimpanzees, strongly supporting the idea that these are two similar but independently evolved locomotor complexes. They go on to argue that humans therefore did not evolve from a knuckle walking ancestor.
These authors have focused on significant differences in the carpal bones that underlie the two forms of knuckle walking. There is a long and heated history of publications in this area (see the Upright Ape[6] for a more detailed discussion of the knuckle walking controversy). Kivell and Schmitt[9] add significant new information about the mechanics and anatomy of knuckle walking as it relates to the very different carpal bone structure in the two lineages. This work does greatly strengthen the argument that there is no proven basis for making these two adaptations into a single shared change that should have been present in the group ancestral to hominins.
On an adaptive basis, one of the arguments most strongly advanced for the preference of closed hand gaits in various hominoids is that the tendons are shortened to result in a passive "hook" of the hand when the arm joints and wrist are extended. This is for the purpose of efficient suspension from branches during hand hanging and locomotion.[14] However, in the light of the Humanian model I have advocated,[6],[7] there would be a multi-million year history of locomotion without hand/arm use and a behavioral reliance on use of the hands for carrying crude tools and small food objects. Closed hand gaits allow hominoids to hold objects in their hands as they locomote - it is easy enough for an adaptationist to explain the adaptive benefit of being able to carry during locomotion when you are in competition with an upright bipedal hominoid. Classical Darwinists often miss this point because of the focus on competition with other species members, but Stephen Jay Gould has pointed out the seemingly obvious importance of competition above the systematic level of the species.[8] This is all about what upright bipeds and knuckle-walkers do on the ground when they compete for resources - and fight with each other - in overlapping territories.
The homeotic mutationist position that I have advocated (The Upright Ape,[6] Homeotic Evolution of the Mammals[5]) tackles the question from the perspective of genetic drive and adaptationist fine tuning. The starting point is the demonstration that the Moroto vertebra demonstrates a truly remarkable transformation of the vertebrate body plan.[5] In most mammals and indeed in most vertebrates - the main dorso-ventral body plane division is placed ventral to the neuraxis, but in a "hominiform" clade that includes hominins, some extant apes, Morotopithecus, Pierolapithecus, Oreopithecus, Orrorin and Sahelanthropus the body plane is shifted to a position dorsal to the neuraxis in the lumbar region.

A number of anatomical transformations in serial homology accompany this change and all of them appear at 21.6 million years ago in Morotopithecus. These include a "box-like" cross section of the lumbar transverse process as opposed to the more standard mammalian flat element and this appears to be evidence of bipedal rather than bimanual orthogrady - e.g. standing and walking rather than arm swinging. The anatomical basis of upright posture in modern humans is very clearly due to this transformation and it is fully present in Morotopithecus. Because of this, I have argued that this feature is the basis of a hominiform clade that separated from the proconsuliform hominoids very soon after the first emergence of hominoids in the early Miocene.

Sudden major transformations of this type are the province of mutationist[17] rather than adaptationist evolutionary progression and are best suited to changes in the major morphogenetic genes such as the homeotic genes. The prime irrefutable example that challenges traditional evolutionists who refuse to understand the implications of modern morphogenetics is the origin of the vertebrates from the invertebrates. Invertebrates have their digestive tract dorsal to their neuraxis, while vertebrates have the neuraxis dorsal to their gut. Invertebrates develop the mouth from the blastopore, but vertebrates develop the anus from the blastopore. To get from the invertebrate body plan to the vertebrate body plan, you have to flip dorsal for ventral and anterior for posterior. An adaptationist argues that there were selective adaptive pressures that gradually turned the proto-vertebrates a few degrees at a time over millions of years. The morphogentic mutationist realizes that this is completely ridiculous and that - as the genes show - the readout gradients in the embryo got flipped.
Vertebrates with the dorsal neuraxis and "aboral" blastopore emerged in abrupt single generation transformations that led to a revolutionary new body plan. In Morotopithecus bishopi we see a similar morphogenetic revolution and the result is the proto-human that does not use its hand or arms to locomote. This most likely occurred in a single generation. A quadrupedal proconsulid-like ape mother and a bipedal orthograde child that was our clade founder once its descendants were isolated by a chromosomal speciation event.
I point out in the Upright Ape[6] that every fossil hominoid species in the hominiform group for which we have post-cranial fossil evidence appears to be an upright adapted form and that there is no evidence yet of any knuckle walkers. The accepted position has been that humans evolved from knuckle walkers around 5-6 million years ago even though there has never been any evidence for this at all. The contrary position that I have advocated is supported by all the evidence from all of the five separate genera of hominiform hominoids known with post-cranial evidence from before the chimp-human split, distributed across time from the 21 to 5 mya, yet my position is the one that has been considered radical.
The general dictates of science are that more credence should be given to a theory with a variety of elements of supporting evidence and less credence to a theory that fits expectations but for which evidence has not yet been found. There are numerous objections to the Humanian Model - a blog I wrote on this subject on Anthropology.net drew nearly 20,000 views and attracted scores of comments making up a good mix of hostile and supportive reactions. However this suggestion has had a very significant impact (Science Daily, blog on afarensis, blog on scientific blogging, MSNBC) because of the strength of the evidence. It was also well received when presented at the 2008 meeting of the American Association of Physical Anthropology[7] and was addressed specifically in presentations others - who reached conclusions consistent with the components of the Humanian Model.[15] David Pilbeam - past Chairman of Biological Anthropology at Harvard - wrote the foreword for the Upright Ape.[6]
Although I first laid out the case in 1986 in Axial Character Seriation in Mammals (republished in 2007),[3] the publication of my 2007 books and papers coincided with the publication of work by Thorpe[19] and Crompton[2] arguing for a upright climbing antecedent for our terrestrial bipedal lineage. There have been three more new hominoid species reported since mid-2007,[18],[12],[10] but none of these finds has included significant new post-cranial fossils. However, analysis of spinal evolution by Rosenman and Lovejoy[15],[16] is consistent with the Humanian model, an analysis of hand bones from Hispanopithecus laeitanus has demonstrated no sign of knuckle walking,[1] and now the new report from Kivell and Schmitt[9] on knuckle walking adds yet more weight to the non-Troglodytian models (no knuckle walking for the human ancestor).

Various studies show the inefficiency of chimpanzee bipedal walking and argue that only by a series of modifications could we arrive at the efficiency of the bipedalism of extant humans.[13] However, this assumes that the common ancestor of chimpanzees and humans walked bipedally the way a modern chimpanzee does when a more correct scenario is probably that the common ancestor walked more as we do - and that its ancestors already had a 15 million year career of minor adaptations improving the effectiveness of bipedal gait.
In the Upright Ape,[6] as well as in the Homeotic Evolution of Mammals[5] and a paper on later Emergence and Optimization of Upright Posture for bipedalism,[4] I update the original (1986)[3] hypothesis by pointing out that the best way to view the locomotor specializations of modern orangs, chimps and gorillas is as examples of "diagonograde" posture. The basic hominiform spine - due to a homeotic mutation - is defective in that it loses the basic mechanical structure that supports the horizontal or pronograde spine in most mammals. This is what occurred in Morotopithecus and has never been restored. In most hominiforms, we retain a very flexible lumbar spine with five or six lumbar vertebrae. However in the orang, the chimp and the gorilla, the lumbar spine is dramatically shortened - to as few as three lumbar vertebrae and independently evolved bony stops that limit hyperextension of the non-orthograde spine appear - each quite independent and different in those three lineages.
Russell Tuttle[20],[21] showed that Orangutans do not knuckle walk - they fist walk. Owen J. Lewis[11] showed that there was no clear shared anatomical basis for knuckle walking in the hominoid wrist (see Upright Ape for numerous references). Kivell and Schmitt[9] have now showed that there are indeed two very distinct functional anatomical patterns for knuckle walking that distinguish chimps and gorillas.
So...if a series of hominiform hominoids share the long flexible lumbar spine that can't function usefully in pronograde or diagonograde postures, if all of these genera of hominiforms demonstrate anatomical features for support upon pelvis below rather than by suspension, and if the orangs, chimps and gorillas have independently evolved short stiff lumbar spines and closed fist diagonograde gaits, then what stands against a Humanian model of human evolution?[6],[7] Certainly there is a mountain of writing, publications, textbooks, theory and belief against it - but there is no actual scientific evidence to support anything other than an upright ancestor for the extant apes and humans.
I have further tried to look closely at what we mean by the word human (OUP blog: Redefining the Word "Human" - Do Some Apes Have Human Ancestors?). As I detail in the Upright Ape, we have Sherwood Washburn to thank for our anthropological definition of the word human and it is basically a hominoid that is bipedal and upright. Clearly, when we look at that century in the early Pliocene that immediately followed the speciation event that sealed the chimp-human split we would now have "humans." These humans would have a brain size no different from a chimp, tool use no different from a chimp, and communication no different from a chimp. The assumption has been that the common ancestor of chimps and humans would look like a chimp - an obvious "ape" - however, what if the common ancestor looked pretty much like the early humans. What if it is the human lineage that retains the primitive traditional hominiform body plan that has been around since the time of Morotopithecus?

I think we all know what we mean by "early human" - they have our basic body form but have smaller ape-like brains, shorter legs, a bigger set of jaws and teeth, and a big toe that spreads out further from the other toes than ours does now. Since the Oxford English Dictionary is probably the best forum for this question, it is on the OUP blog that I placed my piece on redefining the word "human".
From the point of view of cladistics, we really can't use the term "apes" anyhow since this does not form a monophyletic group. From that perspective, we would have to include humans as apes. In fact when you look at the actual sequence of events among the hominiforms and extant apes, "humans" came first, but then the cladists would have us calling the extant apes "humans" and that is not what anyone means. Hominiforms is a good cladistic term for the humans and extant apes. The word human - including early humans and proto-humans - is a good term under Ernst Mayr's view of biological naming in systematics (see discussion in the Upright Ape). We can also say Hominiforms of human body plan (HHBP). All of these issues arise because systematic diversification (speciation) does not necessarily coincide with morphological diversification - some primitive body plans are more effective than the modifications that appear in some descendants.
This is the specific point that Ernst Mayr attacked most specifically in challenging the Hennigian paradigm. In cladistics, whenever a new species arises, we have two daughter species and we say that the parent species is now extinct. Mayr felt that the typical situation was that you had a large slowly evolving population that occasionally emits a speciated branch with a subset of the genetic variability of the parent species. He felt it was biological nonsense to say that a species was now extinct if the genetic makeup of the parent species and the major daughter species were identical.[22] In our situation - we would have a large parent species of human form out of which a small group becomes isolated that leads to the chimpanzees. We agree that the post-split group that is bipedal is now "human" but the pre-split group (from the century before the speciation event) is effectively identical. That pre-split group should be called "human" as well and then we have a human group ancestral to an extant ape lineage - at least in that situation.
One thing that all the naming and systematics logic does not affect is the apparent fact that the immediate ancestors of the all the extant clades of apes & humans had the appearance of modern humans in their body plan.
So we have hominoids that share a modified Y-5 molar and which includes the pronconsuliforms and the hominiforms. The hominiforms are basically upright and of human body form - or humanoid. From the early Miocene to the present, there has always been a lineage of upright primarily bipedal hominiform hominoids whose body plan is of human aspect. In essence these are creatures that don't use their hands and arms for locomotion.
Across time, various groups branch off that are more suited to ecologies and locomotor arenas where humans don't compete as effectively - that is because it has always been difficult for nonhuman-like hominoids to compete against those with human form. The proconsuliforms long since went extinct and the others have not succeeded sufficiently to leave any post-cranial fossils. Meanwhile, the upright bipedal hominiforms with a body plan of human aspect have left an increasingly abundant fossil record that spans 21 million years. They were not stooping forward and they may have been seen occasionally in trees (see my video of Siamang bipedalism and human brachiation: Hominiform Progression on You Tube, in high res on Vuze or as an iTunes podcast). At any point during this time span, if you were to encounter a hominoid it would most likely have been an upright bipedal species that when seen from a distance looked...well... human.
Kivell, T., & Schmitt, D. (2009). Independent evolution of knuckle-walking in African apes shows that humans did not evolve from a knuckle-walking ancestor Proceedings of the National Academy of Sciences DOI: 10.1073/pnas.0901280106
References:
1. Almécija S, Alba DM, Moya-Sola S, Köhler M. Orang-like manual adaptations in the fossil hominoid Hispanopithecus laietanus: first steps towards great ape suspensory behaviors. ProcRoySocB 274:2375-2384 (2007) Open Access
2. Crompton RH, Vereecke EE, Thorpe SKS. Locomotion and posture from the common hominoid ancestor to fully modern hominins, with special reference to the last common panin/hominin ancestor. J Anat 212(4):501-543 (2008) Open Access
3. Filler AG. Axial Character Seriation in Mammals: An Historical and Morphological Exploration of the Origin, Development, Use, and Current Collapse of the Homology Paradigm. Brown Walker Press, Boca Raton, FL, (2007).
4. Filler AG. The emergence and optimization of upright posture among hominiform hominoids and the evolutionary pathophysiology of back pain. Neurosurgical Focus 23(1):E4 (2007).
5. Filler AG. Homeotic evolution in the Mammalia: Diversification of therian axial seriation and the morphogenetic basis of human origins. PLoS ONE 10(e1019) (2007). Open Access
6. Filler AG. The Upright Ape: A New Origin of the Species New Page Books, New Jersey, July (2007).
7. Filler AG. A Humanian Model of Human Evolution: Evidence that habitual upright bipedality is a synapomorphy that defines a hominiform clade of hominoids including humans and all extant apes. American Association of Physical Anthropology, 77th Annual Meeting, Columbus, Ohio April 10, 2008. AJPA 135 (S46): p.96 (2008)
8. Gould SJ. The Structure of Evolutionary Theory. Cambridge, Mass., Belknap Press of Harvard University Press (2002).
9. Kivell TL, Schmitt D. Independent evolution of knuckle-walking in African apes shows that humans did not evolve from a knuckle-walking ancestor. PNAS (2009). paywall
10. Kunimatsu Y et al. A new Late Miocene great ape form Kenya and its implications for the origins of African great apes and humans. PNAS (104) 19220-19225 (2007). Open Access
11. Lewis OJ. Functional Morphology of the Evolving Hand and Foot. Oxford/New York, Clarendon Press ;Oxford University Press. (1989).
12. Pickford M, Coppens Y, Senut B, Morales J, Braga J. Late Miocene hominoid from Niger. C.R. Palevol 8:413-325 (2009). paywall
13. Pontzer H, Raichlen DA, Sockol MD. The metabolic cost of walking in humans, chimpanzees, and early hominins. J Hum Evol 56:43-54(2009).
14. Preuschoft H. Mechanisms for the acquisition of habitual bipedality: are there biomechanical reasons for the acquisition of upright bipedal posture? J Anat 204(5): 363-84. (2004). Open Access
15. Rosenman BA, Lovejoy CO. Developmental anatomy of the hominoid lumbar transverse process: a comparative anatomical framework for examining lumbar natural history in early hominids. American Association of Physical Anthropology, 77th Annual Meeting, Columbus, Ohio April 12, 2008. AJPA 135 (S46): p.183 (2008).
16. Rosenman BA, Lovejoy CO. Relative lumbosacral transverse process length in extant hominoids and australopithecines. American Association of Physical Anthropology, 78th Annual Meeting, Chicago, IL April 3, 2009. AJPA 138 (S48): p.183 (2009).
17. Stoltzfus A. Mutationism and the dual causation of evolutionary change. Evol Dev 8: 304–317. (2006). paywall
18. Suwa G, et al. A new species of great ape from the late Miocene epoch in Ethiopia. Nature 448: 921-924 (2007). paywall
19. Thorpe SKS, Holder RL, Crompton RH. Origin of human bipedalism as an adaptation for locomotion on flexible branches. Science 316: 1328-1331 (2007). requires free registration
20. Tuttle RH . Knuckle-walking and knuckle-walkers: A commentary on some recent perspectives on hominoid evolution. Primate Functional Morphology and Evolution. R. Tuttle. Chicago, Mouton/Aldine: 203-209. (1975).
21. Tuttle RH. Knuckle-walking and the problem of human origins. Science 166(908): 953-61 (1969). paywall
22. Wheeler Q, Meier R. Species Concepts and Phylogenetic Theory : A Debate. New York, Columbia University Press. (2000). Read more!
Thursday, August 13, 2009
A Big Boost for a Revolutionary Theory
One of my more favorite reads was Aaron Filler's Upright Ape: A New Origin of the Species[3], shortly after it was published. The suggestion(s) regarding human evolution were attractive and revolutionary, while the discussions of homeotic mutations and their mechanisms (based upon peer-reviewed work[4]) were enlightening and form part of the foundation of my own approach to understanding how mutation, development, and Darwinian selection work together (a subject I haven't blogged much about although it underlies much of my writing here).
Filler's primary suggestion is that the main stem lineage of the Great Apes has been walking upright since shortly after it diverged from the ancestors of the ancestors of the gibbons and siamangs due to a homeotic mutation that modified the spine to give it the potential to support an upright stance for long periods, something the gibbons and siamangs lack. (Although they can and do walk upright, both in the canopy and on the ground.) One of the implications of this theory is that the common ancestor of humans, gorillas, chimpanzees, and bonobos walked upright, and that knuckle-walking behavior evolved independently in the lineage leading to gorillas from that leading to chimpanzees and bonobos.

Figure 1: Example of the detailed anatomical discussion of the proposed homeotic mutation that lead to the human/great ape backbone structure. Click on image to see illustration and caption at the book's website. (From Reference 3, figure 9-5.)
Now, in the advance online Early Edition of PNAS, we find Independent evolution of knuckle-walking in African apes shows that humans did not evolve from a knuckle-walking ancestor[2] (by Tracy L. Kivell and Daniel Schmitt), unfortunately behind a paywall, that describes research thoroughly undermining the notion that humans evolved from a knuckle-walking ancestor. Let's look at the research first: ...
What Kivell and Schmitt did was examine the "features most commonly thought to reflect knucklewalking behavior in the African ape wrist", in terms of their occurrence and development in many primates, and especially gorillas as well as chimpanzees and bonobos. They found "(i) that most gorillas lack key features that have been assumed to be critical for limiting extension of the wrist during knuckle-walking ([refs]), and (ii) these features are found in monkeys that use a variety of different hand postures and substrates."
Assuming that such features are necessary (or highly desirable) for knuckle-walking, we should find them emphasized in gorillas, and at earlier ages, since gorillas are larger, thus putting more stress on the bones of their hands during knuckle-walking, and normally start earlier during development. Instead:
In addition, the ontogenetic analysis shows that the features of the scaphoid that are assumed to be essential for knuckle-walking ([refs]) are not only inconsistently developed in Gorilla, but, when present, do not appear relatively earlier in development in gorillas (Table 2). Therefore, using the traditional functional interpretation of these features ([ref]), it would appear that the Gorilla radiocarpal joint may be actually less, rather than more, stable in extension throughout ontogeny compared to Pan.The mention of stability during extension of the radiocarpal joint (bending back the wrist) leads to the next important feature of this report: the proposal that the method of knuckle-walking used by gorillas is different from that of chimpanzees and bonobos.
We propose that Gorilla uses a relative ‘‘columnar’’ forelimb posture during knuckle-walking in which the hand and wrist joints are aligned in a relatively straight, neutral posture compared to the more extended postures adopted by Pan (Fig. 3). Animals using a relatively columnar wrist and hand posture would have carpal joints that are in line with the hand and forearm, similar to limb joint position in large graviportal animals such as elephants ([ref]). Supporting loads directly over more vertically-oriented forelimb joints during support phase explains the absence of posited bony adaptations to bending loads in gorillas and also permits more mobility at the joint ([ref]). By contrast, Pan, which exhibit extended wrist postures (Fig. 3), will experience higher bending loads. Thus Pan carpal bones have relatively prominent osteological features that have traditionally, but mistakenly, been interpreted simply as features associated with knuckle-walking in general rather than with a specific posture. The notion that Pan and Gorilla use different wrist postures is consistent with the morphometric data presented here. Although this hypothesis has yet to be explored in detail with videographic data, this idea is further supported by previous research showing that Gorilla exhibits increased wrist mobility compared to Pan ([ref]), a more hyperextended elbow joint ([ref]) and relatively equal length of rays 2 through 4, which creates a larger, more stable area over which to disperse axial loads ([ref]).Imagine the difference between doing push-ups on your fists and on a steel bar you've got your hand wrapped around with your wrist bent back. This is sort of what the difference is, but only sort of. Unfortunately, their "figure 3" is behind a paywall, but the important takeaway is that there is a difference in mechanism, the details of that difference, while important, aren't necessary to the wider implications.
Kivell and Schmitt have provided very strong support for the idea that knuckle-walking works differently, is used differently, and probably evolved independently in these two lineages. A less technically oriented discussion (than the paper) is provided by Science Daily[1], I'm not going to try to replicate their work. (afarensis, FCD is not really convinced, I've discussed Filler's theory in comments on his (old) blog, and I've expanded and modified some of my arguments there in this post.)
Support for the Upright Ape
Instead, I'm going to discuss how this new research relates to the "Upright Ape" hypothesis. One key hypothesis here is that the homeotic mutation to the spine "pre-adapted" it to support the upper body when standing, walking, or running upright. I won't go into the details, but with this adaptation in place, we need to ask precisely why any possessor would go in for knuckle-walking. The answer is that the requirements of brachiation (swinging from the arms) include suppressing a twisting movement between the hips and the shoulders, a need that grows greater with greater size (mass). An ancestral "Great Ape" weighing no more than a typical gibbon might have found this movement easy to suppress, but a larger ape that depended on brachiation would have been under some selective pressure to suppress this movement.
(Another possibility, IMO, is that the lumbar structure of gibbons has actually become adapted to use the springy structures of the lumbar transverse process (LTP) to provide energy storage from/to this twising moment, which in Great apes must be suppressed by muscular action at the cost of energy. AFAIK no research has been done into this possibility. I'll mention that the springy structures involved are used in most mammals to counter the drop of the belly, allowing it to be suspended during walking and storing energy during running/leaping, while in Great apes it tends to counter the forward droop of the upper body while standing upright (because it has been moved dorsally of the center of flexion (bending) of the backbone by the homeotic mutation). For more detail, I must refer you to the book,[3] I just don't have the expertise, and haven't done the detailed research, to attempt it. Thus I must also leave my suggestion regarding the gibbons as a possibility, until I find research (if any) into the details of how these structures work during active brachiation.)
Filler makes the case that the long hip structures and functionally short lumbar structures in African Great Apes represent adaptations to this selective pressure, and then goes on to argue that there are enough differences that this adaptation was probably made independently in chimpanzees/bonobos vs. gorillas. He also makes a persuasive case that this adaptation is responsible for making it much harder for African apes to walk upright efficiently, because they are forced to twist their body with, rather than counter to, their hips. This, then, would explain the parallel adoption of knuckle-walking by the two lineages during or after their divergence from each other and also from that leading to humans, which appear to have happened in one event (with gorillas diverging earlier from a lineage that later split to create those leading to chimps/bonobos on the one hand and humans on the other, although IMO a more complex triple split with a sustained period of reduced gene flow is more consistent with the evidence). The common ancestor was either small, made little use of brachiation, or both. After the split, both lineages developed into comparatively large-bodied brachiators, later becoming increasingly terrestrial and now dependent on knuckle-walking where the common ancestor would have used upright walking somewhat similar to that of gibbons and siamangs.
A suggestion (by Thorpe, Holder, and Crompton) that bipedalism developed among arboreal apes as "an Adaptation for Locomotion on Flexible Branches"[7] was countered by a comment (by Begun, Richmond, and Strait) referring to the evidence of shared features indicative of knuckle-walking and appealing to parsimony as demonstrating its use in the common ancestor of humans and African apes.[9] With this paper,[2] we see the argument of the Begun team dissolve: There is no longer any strong evidence of knuckle-walking in the common ancestor, or any of the central lineage(s) of the great apes.
The Thorpe/Crompton team hasn't stood still, a more recent review examines a great deal of detail regarding the mechanics of locomotion, especially with respect to foot-ground contact and the movement of the center of gravity (COG), in support of an arboreal origin for bipedalism. They deal with the knuckle-walking hypothesis thus:[5]
To our knowledge, Gebo is the only supporter of the knuckle-walking hypothesis to have offered a functional argument linking knuckle-walking and the origins of modern human bipedality, but as we have shown, his argument, which is restricted to foot function, is poorly supported by the data. No explanation appears to have been offered of how the characteristic flexed hindlimb postures of knuckle-walking gait could have been pre-adaptive for the extended hindlimb postures of modern human walking, or indeed how the transformation could have occurred. Further, the knuckle-walking hypothesis requires that orthogrady evolved twice: from pronograde simians to the orthograde crown hominoids – and then following an adoption of terrestrial knuckle-walking by the common African ape ancestor to orthograde bipedality in hominins. We prefer the alternative, that knuckle-walking evolved evolved independently in gorillines and panins, as the ontogeny of phalangeal and metacarpal scaling ([ref]) and of other 'knuckle-walking features' ([ref]) is different in panins and gorillines, suggesting that knuckle-walking evolved more than once among the African apes.Kivell and Schmitt have certainly provided great support for this preference. However, there are some problems with this latest (AFAIK) volley.
Although they mention Filler's work with homeotic mutations,[4] Crompton et al. do not (as far as I can tell) take note of the implications of lumbar stiffening as an adaptation for brachiation in larger-bodied apes, although it seems fully consistent with their model. Apes that spent most of their time clambering around in an upright position (with little brachiation) might well have had no need for such lumbar stiffening. More importantly, Crompton et al. fail to allow for the selective advantage of the springy backbone supporting the upper body. (In fact, even Filler puts more emphasis on support standing or walking rather than running.)
Crompton et al. actually discuss the use of heel-strike in walking by various apes, and mention the "lack of heel-strike in modern hylobatids",[5] but fail to make the connection to the shock-absorbing, and especially energy-storing capacity, of the springy lumbar structures of Great apes (including humans). Of course, I don't recall any special emphasis on this in the book, but then the Filler wasn't arguing against the Thorpe/Crompton suggestion of arboreal bipedalism. Although I can't speak as an expert, it seems to me that the advantages of the Great Ape lumbar structure would primarily apply to ground and very large branches/trunks, where the elastic support for the upper body could cushion the jarring effect of the heel-strike and store at least part of its energy. Landing on smaller branches with a heel-strike would be less problem, as they could provide the springy support, alternatively too much force could destabilize things, so the heel-strikes used by Great Apes on small branches are probably as careful as those of humans on icy pavement.
Problems with Current Selective Models
What this suggests to me is that the stem lineages of great apes relied much more on terrestrial movement than Crompton et al. suggest.[5] Of course, reliance doesn't necessarily mean frequent use. One major problem I have with all these studies is their tacit assumption that energy efficiency, or frequent use, represent a good measure of selective advantage. Apes are social animals, which means that intra-specific competition is extrememly important in selection, both within and between social groups. The "winners" of these competitions could be expected to have as much as they needed to eat, so energy efficiency would be less important than victory. Similarly, all such competition is ultimately "life-and-death", at least reproductively, given the reproductive advantages accruing to high-status individuals, so adaptations that reduce the risk of high-risk behavior involved in such competition would likely be selectively advantageous even at the cost of efficiency, and even when the behavior involved is not frequent.
In arguing for an arboreal origin for bipedalism, Crompton et al. state:
Kingdon (2003) has proposed that in Africa, from the Middle to Late Miocene, fragmentation of closed forest alternated with reclosure and reinvasion of gallery-forest, moist-woodland and rainforest environments, changes also clearly documented by Elton (2008)[13]. Within branches of both the Pan and the Gorilla lineages, the height-range and frequency of vertical climbing locomotion must have increased, suggests Kingdon (2003), to facilitate access to preferred foods in the main and emergent canopy, while permitting travel between trees in broken-canopy woodland on the ground. Middle Miocene crown hominoids, such as Hispanopithecus (Dryopithecus) laietanus, are of a similar size range to living great apes, although undoubtedly male body weights for living great apes exceed estimated values for the Miocene. Thorpe et al. (2007a)[7] thus speculate that the gorillines and panins, independently, became increasingly specialized on forelimb power, to sustain safety and effectiveness of increased vertical climbing, and so tended to adopt similar, extended-elbow, flexed-hip-and-knee kinematics when moving on the ground, hindlimb musculature being unable readily to sustain hip and knee extension and hence a bipedal gait. Hominin ancestors, we speculate, sacrificed continued access to the canopy, and became increasingly ground and small-tree dwellers. Lacking the increased specializations for vertical climbing, they were able to sustain a bipedal gait – and we may add, inasmuch as they were terrestrial, that they could sacrifice coronal-plane mobility, which also characterizes the orangutan, in favour of more effective parasagittal force-production.Alternatively, my best interpretation of Filler would suggest that the "gorillines and panins, independently, became increasingly specialized on" brachiation, perhaps not often but in selectively critical circumstances, producing the independent lengthening of the hip and stiffening of the lumbar region adduced by Filler, but making the ancestral bipedalism very inefficient, producing independent adaptations for knuckle-walking. "Hominin ancestors", pretty much retained the ancestral condition, which would be very Australopithecus-like. Only with the appearance of the lineage(s) called "Homo habilis" and the transition to a Homo erectus-like form do we find the more sophisticated gaits found in modern humans.
The suggestion that the ancestral "Great Apes" possessed a functional upright bipedalism pretty much identical to that of the Australopithecenes, with all the modern "Great Ape" lineages representing separate offshoots independently adapted for arboreal lifestyles at the expense of the ancestral bipedalism, is generally consistent with both Filler and the Thorpe/Crompton team. However, the issue of the relationship between heel-strike and the lumbar modifications speaks for a much greater reliance on terrestrial movement than is consistent with the Thorpe/Crompton arboreal explanations.
Although Filler didn't speak to it, I think we should consider the environmental implications: conditions were not always a closed canopy:
Mid and late Miocene African catarrhines are associated with a variety of habitats, over time as well as in different places ([refs]). Early Miocene sites such as Mfwangano Island and Songhor, sampling several catarrhine genera including Proconsul and Rangwapithecus, probably had environments containing evergreen, multi-canopied forest ([refs]). The early Miocene catarrhines radiated extensively into such arboreal habitats, filling frugivorous, folivorous, above-branch and mixed arboreal/terrestrial niches ([ref]). The slightly more recent (c. 18 Ma) Hiwegi Formation on Rusinga Island, by contrast, appears to have single-canopied, disturbed woodland, known best from a rich floral assemblage ([ref]). Unfortunately, it has no directly associated ape fossils although several taxa are known from Rusinga Island deposits of a similar age ([ref]).[13]Given the widespread lack of fossils, we can't assume that any area lacking fossils lacked some sort of ape population. Proconsul, according to Filler,[3] did not possess the specialized lumbar structures found in Great Apes, although a very similar species did. The "evergreen, multi-canopied forest" mentioned by Elton[13] may well have been similar to the primary environments of both African and Asian modern Great Apes, while the "single-canopied, disturbed woodland" is another type of environment entirely. With broken canopies, and even a single canopy which would not have been completely opaque, there would have been much more undergrowth. This in turn would have led to much more scrambling and clambering moving over the ground. The "disturbed" aspect of the woodland suggests considerable downwood, placing all sorts of obstacles to overland transport. I would suggest that from the earliest expansion of the "Great Apes", that is those possessing the lumbar adaptations adduced by Filler, they specialized in rapid movement through this sort of terrain.

Figure 2: Example of "disturbance": damages of storm Kyrill in Wittgenstein, Germany. (From Wiki.)
In areas with high relief (from erosion), there wouldn't have been a clear boundary between this sort of movement and low arboreal movement through the "single-canopied, disturbed woodland". Mammals the size of Proconsul spend a lot more energy in vertical movement than horizontal,A1 so an ability to move horizontally, or as close to it as possible, through a variety of micro-landscapes would be adaptive. So would an ability to run (when necessary, see above) over hard ground and large branches/logs at similar levels. Unlike the "assisted bipedal walk"[12] that may be the safest way to cross high bridges, bipedal running during necessary high-risk activity would probably have depended on balance. This is hardly surprising, many primates can be taught to ride a bicycle, a task demanding great balance.

Figure 3: High relief terrain, from Oregon. This is badlands, much less wooded than the type of terrain I'm discussing, but it allows you to see the relief because of the lack of vegetation. Imagine forest over this terrain, with frequent disturbance as in Figure 2. (From the files of OregonLive.com.)
Given the generally Australopithecus-like fossils being found in Africa, some evidently dating back as early, or earlier, than the best date for the split between the lineages leading to humans vs. chimps/bonobos,[5] [13] even parsimony, unreliable as it is, would suggest that the common ancestor was pretty much Australopithecus-like.
We can extend this argument to orangutans: given the many apparent similarities shared by humans and orangutans but not by chimps or gorillas, these may well have been inherited from an Australopithecus-like ancestor. In this case the arboreal (brachiating) adaptations of all three lineages would have been evolved independently, with different suites of ancestral features retained to be shared with humans.
Personally, I find Filler's hypothesis very convincing, more so than either the Knuckle-walking or the arboreal bipedalism explanations. And this paper provides strong support (although it doesn't say anything against arboreal bipedalism).
Kivell, T., & Schmitt, D. (2009). Independent evolution of knuckle-walking in African apes shows that humans did not evolve from a knuckle-walking ancestor Proceedings of the National Academy of Sciences DOI: 10.1073/pnas.0901280106
Appendix: You can use the back key to return to where you were, or click on the quoted text to return to the line with the footnote.
A1. "Mammals the size of Proconsul spend a lot more energy in vertical movement than horizontal": This is because the mammalian ratio of basal metabolic rate to weight decreases with weight differently than the energy involved in vertical movement varies with weight, as does the energy involved in movement at all. I have been unable to find any discussion of this online, and can't even recall where I read it. I find it hard to believe that this subject has been ignored in discussions of arborial vs. terrestrial movement: I suspect there's something wrong with my search technique.
Basically, an animal the size of a squirrel spends almost as much energy moving horizontally as vertically, and the difference isn't all that large (for mammals with their high metabolic rate) from standing still. Both differences increase with increasing body weight (technically, mass). Obviously, the effect also varies by type of movement.
Links: I've included only the links called out in this leader. Not all of these links are called out in the text. Use the back key if you came via clicking a footnote.
1. Bipedal Humans Came Down From The Trees, Not Up From The Ground ScienceDaily (Aug. 11, 2009) not peer reviewed
2. Independent evolution of knuckle-walking in African apes shows that humans did not evolve from a knuckle-walking ancestor paywall
3. The Upright Ape: A New Origin of the Species by Aaron G. Filler MD, PhD
4. Homeotic Evolution in the Mammalia: Diversification of Therian Axial Seriation and the Morphogenetic Basis of Human Origins open access
5. Locomotion and posture from the common hominoid ancestor to fully modern hominins, with special reference to the last common panin/hominin ancestor paywall
6. Orangutan positional behavior and the nature of arboreal locomotion in Hominoidea
7. Origin of human bipedalism as an adaptation for locomotion on flexible branches free registration required
8. The Origins of Human Bipedalism paywall
9. Comment on "Origin of Human Bipedalism As an Adaptation for Locomotion on Flexible Branches" Open Access
10. Response to Comment on "Origin of Human Bipedalism As an Adaptation for Locomotion on Flexible Branches" Open Access
11. Locomotor Ecology of Wild Orangutans (Pongo pygmaeus abelii) in the Gunung Leuser Ecosystem, Sumatra, Indonesia: A Multivariate Analysis Using Log-Linear Modelling paywall
12. Orangutan Positional Behavior and the Nature of Arboreal Locomotion in Hominoidea
13. The environmental context of human evolutionary history in Eurasia and Africa Open Access
Read more!
Subscribe to:
Posts (Atom)